KAPITEL 6. Frugtvækst og frugtkvalitet
Index
6.2. Frugtvækst og -udvikling på basis af source-sink-relationer
6.2.1. Frugt/blad-forhold (blad/frugt-forhold)
6.2.2. Source-aktivitet
6.2.3. Sink-aktivitet
6.2.4. Relativ sink-styrke i andre organer
6.3. Blad/frugt-forhold eller frugt/blad-forhold hos æble
6.4. Source-aktivitet (bladaktivitet). Æble
6.6. Vekselvirkning mellem source-sink-faktorer (æble)
6.7. Blomsterkvalitet, fysiologisk kvalitet af unge frugter
6.8. Frugtstørrelsesvariation inden for et træ (æble)
6.9. Frugtvækst og -kvalitet hos blommer
6.10. Frugtvækst og frugtkvalitet hos kirsebær
6.11. Frugtvækst og frugtkvalitet hos solbær (Ribes)
6.12. Frugtvækst og frugtkvalitet hos jordbær
6.12.1. Blomsterkvalitet (antal æganlæg pr. blomst)
6.12.2. Forholdet achener/æganlæg
6.12.3. Bærvækst, bærsvulmning pr. achene
6.12.4. Bærkvalitet
Frugttilvæksten bestemmer frugtens størrelse og form. Frugtstørrelsen er en vigtig udbyttekomponent og for konsumfrugt også en vigtig kvalitetskomponent. Udvikling af farve kan også være en vigtig kvalitetsfaktor. Forskellige indre kvalitetskomponenter som fasthed, konsistens, saftighed, sødhed, surhed, aroma, vitaminer og indhold af fremmedstoffer har betydning for nydelses-, smags-, forarbejdnings- og ernæringsværdien. Særlige grupper af indholdsstoffer som sukkerarter, organiske syrer, aromastoffer, farvestoffer og vitaminer er afgørende for nogle af de omtalte kvalitetskomponenter.
Der hersker en stor variation med hensyn til såvel ydre som indre kvalitetsfaktorer. I det følgende gives der først en oversigt over den genetiske variation. Derpå behandles mere udførligt fysiologiske aspekter vedrørende frugtvækst og frugtudvikling som baggrund for dyrkningstekniske, klimatiske og andre påvirkninger. Den udviklingsmæssige påvirkning i relation til frugtmodning samt vitaminer, aroma og farvestoffer, som er særligt knyttet til modningsprocesserne, behandles senere i separate kapitler.
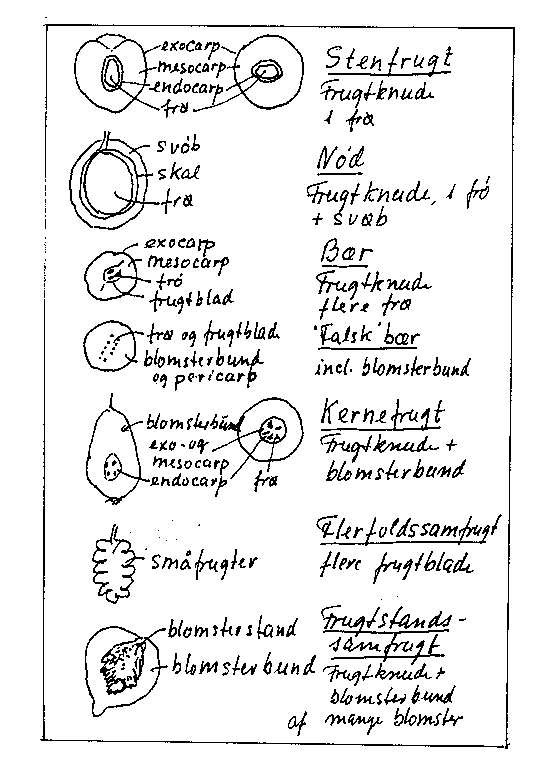
Såvel ydre som indre frugtegenskaber varierer stærkt mellem frugtarterne. Botanisk er der forskellige frugttyper, som vist i bilag 6-1. Der kan være tale om 1-frøede frugter udviklet alene fra frugtknuden (frugtblade, som kan opdeles i endocarp, mesocarp og exocarp). Stenfrugter har en kødet mesocarp (Prunus-arterne), mens nødderne har et tørt ydre (hasselnød). Valnødder og mandler kaldes i en frugtavlsmæssig sammenhæng for nødder, idet de bliver relativt tørre, men er botanisk set stenfrugter. Kødede, flerfrøede frugter bestående af frugtknuden er bær (Ribes, vindrue). Dele af blomsterbunden kan indgå som hos blåbær ("falsk" bær). Kernefrugter er også "falske" i botanisk henseende, idet de består af frugtknude + blomsterbund udviklet af oversædige blomster. Flere frugtknuder indgår og falder af samlet hos flerfoldssamfrugter, som hindbær. Hos brombær og jordbær indgår også blomsterbunden. I frugtstandssamfrugter som figen, morbær mv. indgår desuden dele af blomsterstanden.
Bilag 6-1. Frugttyper. Efter Westwood (1978).
Knyttet til de botaniske frugttyper er der også store forskelle i frugtstørrelse og -form. Der er som bekendt også store farveforskelle. Også sammensætningen af frugterne varierer (bilagene 6-2, 6-3). De fleste frugter er vandholdige (ofte over 80% vand). "Nødderne" er dog ret tørre, de fleste af dem indeholder protein og fedt, ligesom det er disse frugter, som indeholder mest B-vitamin samt jern. I de øvrige frugtarter er kulhydrater den dominerende del af tørstoffet. Abrikoser indeholder A-vitamin. C-vitamin findes især i solbær, men også i andre bærfrugter. Vandholdige frugter er relativt kaliumrige.
Bilag 6-2. Størrelsesorden for indholdsstoffer i forskellige frugtarter. Efter forskellige kilder (Westwood 1978; Keipert 1981; Anon. 1984).
Frugtart |
Vand |
Protein 1) |
Fedt 1) |
Kulhydrat |
Domin. Sukker* |
Domin. Syre** |
Vitamin C 2) |
Kalium 2) |
Abrikos |
85 |
1 |
< 1 |
14 |
-- |
æ |
10 |
280 |
Appelsin |
86 |
1 |
< 1 |
10 |
-- |
c |
56 |
-- |
Blomme |
82 |
< 1 |
1 |
16 |
g,s |
c |
5 |
170 |
Blåbær |
84 |
< 1 |
< 1 |
15 |
s |
b |
17 |
70 |
Brombær |
85 |
1 |
< 1 |
11 |
g,f |
æ |
20 |
190 |
Citron |
85 |
< 1 |
< 1 |
8 |
-- |
c |
40 |
-- |
Fersken |
85 |
< 1 |
< 1 |
10 |
g |
æ |
8 |
200 |
Hasselnød |
86 |
13 |
62 |
17 |
-- |
-- |
-- |
700 |
Hindbær |
88 |
1 |
1 |
10 |
f,g,s |
c |
25 |
190 |
Hyldebær |
80 |
3 |
1 |
13 |
-- |
æ |
28 |
300 |
Jordbær |
90 |
< 1 |
< 1 |
8 |
g,f |
c |
63 |
160 |
Kastanie |
52 |
2 |
3 |
41 |
-- |
-- |
0 |
450 |
Kirsebær |
82 |
2 |
< 1 |
14 |
g,f |
c |
12 |
190 |
Kiwi |
84 |
1 |
< 1 |
14 |
-- |
-- |
70 |
-- |
Mandel |
5 |
20 |
54 |
19 |
-- |
-- |
1 |
780 |
Pekannød |
3 |
9 |
72 |
14 |
-- |
-- |
2 |
600 |
Pistacie nød |
6 |
20 |
53 |
19 |
-- |
-- |
4 |
970 |
Pære |
83 |
< 1 |
< 1 |
14 |
f |
æ |
5 |
130 |
Ribs |
86 |
1 |
< 1 |
9 |
g,f |
c |
35 |
240 |
Solbær |
83 |
2 |
< 1 |
12 |
f,g |
c |
180 |
330 |
Stikkelsbær |
90 |
1 |
< 1 |
9 |
g,f |
c,æ |
40 |
180 |
Tomat |
94 |
1 |
< 1 |
4 |
-- |
æ |
20 |
-- |
Tyttebær |
86 |
1 |
1 |
12 |
s |
b |
11 |
70 |
Valnød |
24 |
13 |
58 |
13 |
-- |
-- |
2 |
450 |
Vindrue |
83 |
1 |
< 1 |
16 |
g,f |
v |
5 |
-- |
Æble |
84 |
< 1 |
< 1 |
12 |
f |
æ |
8 |
110 |
1) g pr. 100 g friskvægt 2) mg pr.100 g mg pr.100 g, g = glucose; f = fructose; s = sucrose. ** æ = æblesyre; c = citronsyre; b = benzoesyre; v = vinsyre.
Bilag 6-3. Udbytteniveauer og frugtkarakterer. Gennemsnit og variationsområde i danske sortsforsøg i forskellige frugtarter.
Jordbær |
Solbær |
Ribs |
Hindbær |
Brombær |
Surkirs. |
Æble |
||
t/ha |
gns. |
25 |
8 |
15 |
7 |
2 |
22*** |
-- |
var. |
11-40 |
2-17 |
11-22 |
3-13 |
0-5 |
7-36*** |
-- |
|
g/frugt |
gns. |
12 |
0,9 |
1,1 |
2,6 |
2,9 |
4,5 |
-- |
var. |
6-19 |
0,6-1,4 |
0,9-1,4 |
1,4-3,6 |
2,1-4,4 |
2,7-6,4 |
-- |
|
Total tørstof % |
gns. |
9 |
22 |
17 |
12 |
14 |
-- |
-- |
var. |
7-11 |
17-28 |
15-23 |
10-14 |
12-17 |
-- |
-- |
|
Opløs.tørstof % |
gns. |
7 |
14 |
9 |
9 |
8 |
16 |
12 |
var. |
6-9 |
10-17 |
7-11 |
6-12 |
7-10 |
11-18 |
5-15 |
|
Titrerbar syre g/100 g |
gns. |
1,0 |
3,2 |
3,5 |
2,2 |
1,4 |
1,8 |
1,0 |
var. |
0,8-1,2 |
2,7-4,1 |
2,7-4,1 |
1,5-3,2 |
1,0-1,9 |
1,1-2,3 |
0,4-1,7 |
|
C-vitamin mg/100 g |
gns. |
65 |
163 |
-- |
-- |
6 |
-- |
20 |
var. |
45-99 |
95-253 |
-- |
-- |
4-8 |
-- |
1-59 |
|
Farve, mg/g* |
gns. |
-- |
3,4 |
0,16 |
0,8 |
-- |
1,2 |
-- |
var. |
-- |
2,6-5,0 |
0,12-0,20 |
0,3-1,3 |
-- |
0,3-2,8 |
-- |
|
Kilde** |
1435b |
1892m |
991m |
889m |
1266m |
1413b |
||
1555b |
1745m |
1522m |
1864b |
|||||
1778b |
1553b |
1780m |
1971b |
|||||
926m |
* Som malvidinchlorid.** Henviser til beretninger (b) og meddelelser (m) fra Statens Planteavlsforsøg.*** Ved "fuld" bæring.
Også kulhydratsammensætningen varierer; fructose er ofte den dominerende sukkerart, men det kan også være en af de andre sukkerarter (bilag 6-2). Indholdet af organiske syrer varierer, æble- eller citronsyre er oftest den dominerende syre. Ikke blot indholdet, men også sammensætningen af kulhydrater og syrer har betydning for det smagsmæssige indtryk, idet de forskellige sukkerarter og syrer har forskellig smagsværdi (bilag 6-4). Aromastoffer, vitaminer og farvestoffer behandles nærmere i et følgende kapitel.
Bilag 6-4. Relative værdier for organoleptisk sødhed hos sukkerarter (sukrose = 100) og for surhed hos organiske syrer (1-9; 9 = surest).
Relativ sødhed: |
Relativ surhed: 1-9; |
||
Fruktose |
173 |
Æblesyre |
8,2 |
Sukrose |
100 |
Vinsyre |
4,3 |
Glukose |
74 |
Citronsyre |
3,6 |
Sorbitol |
50 |
Fosforsyre |
1,0 |
Også inden for de enkelte frugtarter er der en betydelig genetisk variation. Dette gælder indre (bilag 6-3) såvel som ydre frugtegenskaber; mange af dem er vigtige mål ved forædling. Med hensyn til den sortsmæssige variation i øvrigt henvises til senere behandling, her skal blot nævnes, at den er særdeles vigtig, og i mange tilfælde overgår den den variation, som dyrkningsteknik og andre faktorer kan forårsage.
6.2. Frugtvækst og -udvikling på basis af source-sink-relationer
Frugttilvæksten kan være afhængig af blomsterkvaliteten (blomsterudviklingen) og dermed den fysiologiske kvalitet af den helt unge frugt; her kan bestøvning og befrugtning også spille en rolle; det gælder især i arter med mange frø pr. frugt (Ribes, Rubus og jordbær).
I kap. 5 er vist, at frugter for en stor del opbygges på grundlag af assimilater, som tilføres fra bladene, og at relationer mellem blade, frugter og andre organer har betydning, ikke blot for assimilatfordelingen, men også for den intensitet hvormed assimilatomsætningen sker. Det er derfor naturligt at forestille sig, at frugtvækst og -udvikling vil påvirkes af assimilatniveau eller assimilat-tilgængelighed i planten, som igen vil være afhængig af forholdet mellem assimilatproducerende organer (blade) og assimilatforbrugende organer (her især frugter) som defineret ved følgende udtryk (Patrick 1976):
Source-styrke / sink-styrke = source-størrelsexsource-aktivitet / sink-størrelsexsink-aktivitet
Source-styrke = assimilatoriske vævs kapacitet med hensyn til syntese af stoffer til eksport.
Source-størrelse = eksempelvis bladareal.
Source-aktivitet = assimilatorisk hastighed.
Sink-styrke = vævs potentielle kapacitet med hensyn til akkumulering af metaboliter.
Sink-størrelse = eksempelvis frugtantal.
Sink-aktivitet = potentiel metabolit-optagelseshastighed pr. sink-enhed og tidsenhed.
Både størrelses- og aktivitetskomponenter kan tænkes påvirkede, og dette vil igen virke ind på assimilatomsætning og frugtvækst som illustreret i bilag 6-5.
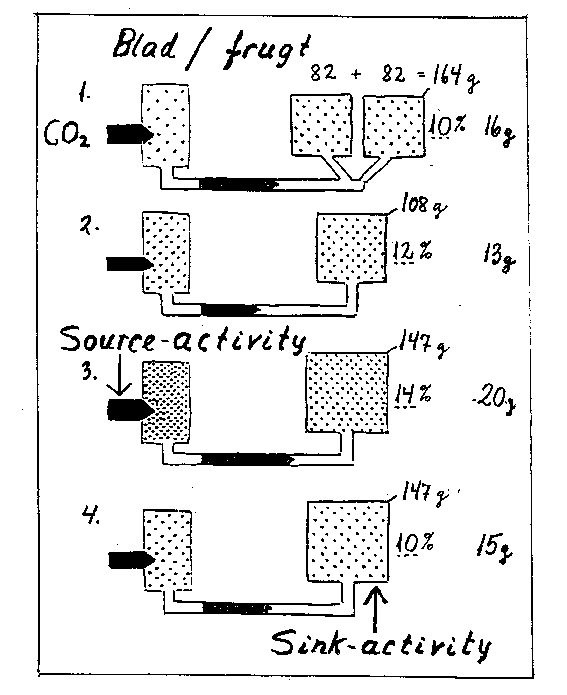
Bilag 6-5. Hypotetisk skitse over source-sink-elementers betydning for assimilatomsætning og frugtudvikling. Venstre dele illustrerer blade, højre dele frugter, hvor tallene angiver hypotetiske værdier for friskvægt, tørstofkoncentration og tørvægt. Se i øvrigt teksten.
6.2.1. Frugt/blad-forhold (blad/frugt-forhold)
Sammenlignes et større frugt/blad-forhold (1, se bilag 6-5) med et mindre (2), vil de flere frugter bevirke et større "træk" i assimilaterne. Derved animeres bladene til en større produktion (større fotosynteseintensitet), ligesom transporten ud af dem går hurtigere. Da der imidlertid ikke sker nogen fuldstændig kompensation, bliver assimilatniveauet lavere. Den enkelte frugt vokser langsommere, samtidig "fortyndes" systemet, så at koncentrationen af visse stoffer i blade og frugter bliver lavere; det gælder fx tørstofkoncentrationen i frugten.
Hvis bladenes fotosynteseintensitet øges fx via mere lys (bilag 6-5, 3 kontra 2), vil der blive et større "tryk" fra bladene, assimilattilgængeligheden bliver større, frugterne vokser mere samtidig med, at ophobningen i både blade og frugter øges.
6.2.3. Sink-aktivitet
Mens der ovenfor er skitseret virkninger på assimilattilgængeligheden, og dermed på frugtvæksten, er der her tale om andre faktorer, som påvirker frugtvækst, fx via stoffer tilført fra rødderne. Ved en øget sink-aktivitet i frugterne (bilag 6-5, 4 kontra 2) vil disse "trække" kraftigere i assimilaterne. Måske sker der en vis kompensation ved øget fotosyntese- og transporthastighed, men systemet fortyndes, så frugterne samtidig får et lavere tørstofindhold. Bemærk, at der ved denne virkning vil være en negativ korrelation mellem frugtvækst og koncentrationer af tørstof, modsat en positiv korrelation i de to førstnævnte tilfælde.
6.2.4. Relativ sink-styrke i andre organer
Hvis assimilatforbruget er særligt stort eller særligt lille i andre organer på grund af særlige virkninger på vækstintensiteten eller vækstens varighed her, kan assimilattilgængeligheden for frugterne blive påvirket og dermed frugtvækst og -indhold på lignende vis som i eksemplerne 1-3, bilag 6-5.
I de følgende afsnit er de nævnte situationer nærmere belyst med resultater fra forsøg med æble. Dernæst følger en vurdering af vigtige frugtvækstfaktorer i nogle af de andre frugtarter. Bilag 6-6 viser en oversigt over vigtigheden af de enkelte faktorer i forskellige frugtarter.
Bilag 6-6. Virkninger på frugtvækst og frugtudvikling (+ = positiv korrelation; -- = negativ korrelation).
Frugtstørrelse |
Tørstof (sukker) |
Syre |
Aroma og C-vitamin |
Gul grund- farve |
Rød dæk-farve (æbler) |
|
Blomsterkvalitet1) |
+ |
|||||
Bestøvningseffekt1) |
+ |
|||||
Blad/frugt2) |
+ |
+ |
+ |
+ |
+ |
|
Frugt/blad2) |
-- |
-- |
-- |
-- |
-- |
|
Source-akt. (lys)3) |
+ |
+ |
+ |
+ |
+ |
[+] |
Sink-akt.4) |
+ |
-- |
-- |
|||
Sink-styrke-reduktion i andre organer |
+ |
+ |
||||
Udvikling5) |
+ |
+ |
||||
Modning5) |
+ |
--/0 |
[±] |
[+] |
[+] |
1) Især frugter med mange frø (jordbær, Ribes, Rubus); især med hensyn til inden for plante variation.
2) Virkning via assimilattilgængelighed. Især storfrugtede arter.
3) Formentlig virkning i alle arter, dog nok mest i storfrugtede arter, relativt lidt i jordbær.
4) Virkning i alle arter, vigtigste virkningsfaktor i Ribes.
5) Virkning i alle arter, dog største slutforskelle, hvor frugten har en lang udviklingsperiode.
6.3. Blad/frugt-forhold eller frugt/blad-forhold hos æble
Disse forhold er naturligvis blot to sider af samme sag. Men da de begge anvendes, kan det være nyttigt at se på dem hver for sig. Med stigende blad/frugt-forhold falder bladenes fotosynteseintensitet, mens bladenes indhold af bl.a. stivelse stiger, ligeledes øges frugtvæksten såvel som koncentrationen af total tørstof, opløseligt tørstof (sukker) og syre i frugterne (bilagene 6-7, 6-8). Dette svarer til de postulerede sammenhænge i afsnit 6.2.1. Virkningsgraden kan variere med sorten. Frugternes fasthed kan også øges ved stigende blad/frugt-forhold.
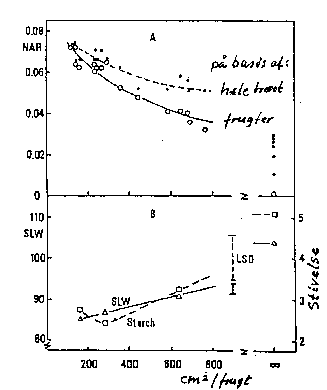
Bilag 6-7. Virkning af blad/frugt-forhold på A) bladenes netto-assimilations-hastighed (NAR) samt B) bladtykkelse (SLW) og stivelsesindhold (Hansen 1982).
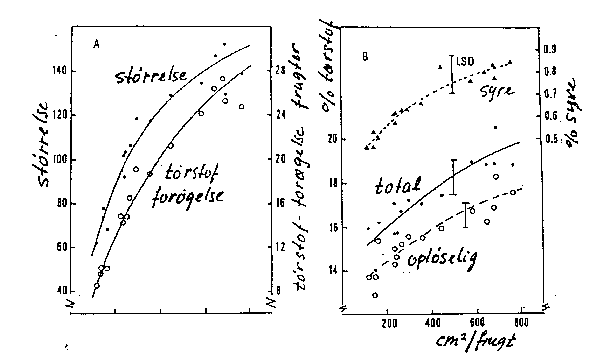
Bilag 6-8. Virkning af blad/frugt-forhold på A) frugtvækst (-størrelse) og på B) indhold af totalt tørstof, opløseligt tørstof og titrerbar syre (Hansen 1982).
Bilag 6-9 viser virkningerne som funktion af frugt/blad-forhold. Disse sammenhænge vil også nogenlunde svare til relationerne mellem frugtstørrelse, frugtudbytte og frugtantal på hektarbasis, hvis vi erstatter bladmængde med jordareal. Under markforhold vil det være praktisk umuligt at bestemme bladmængden. Af kurverne vil fremgå, at udbyttet pr. ha stiger med frugtantallet pr. ha, men med aftagende styrke, mens frugtstørrelse falder, men også med aftagende styrke.
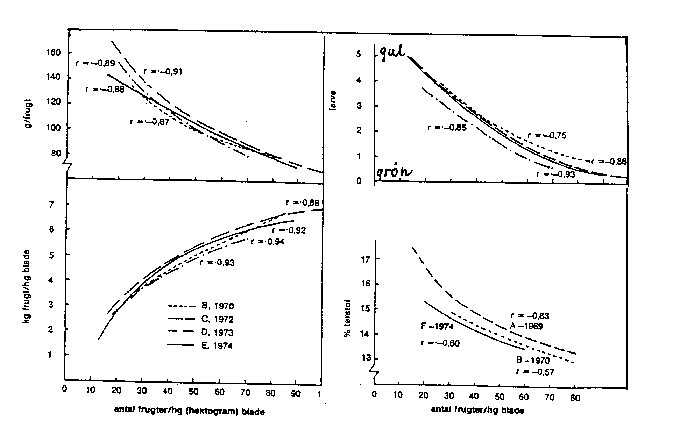
Bilag 6-9. Udbytte og frugtudvikling som funktion af frugt/blad-forhold. 'Golden Delicious'. Efter Hansen (1977).
Frugtudtynding er det vigtigste middel til at påvirke frugt/blad-forholdet. Ændring af bladarealet vil naturligvis også virke, fx ved sommerbeskæring (afsnit 14.3.1).
6.4. Source-aktivitet (bladaktivitet). Æble
Det er velkendt, at fotosynteseintensiteten øges med stigende CO2-koncentration. Samtidig øges transporthastigheden af assimilater ud af bladene, og koncentrationen af visse stoffer i bladene stiger (bilag 6-10).
Bilag 6-10. Virkning af CO2-niveau på sukkerindhold, og på transport ud af blade efter tilførsel af 14CO2 (Hansen 1975).
CO2-niveau |
Reduceret |
Normal |
Øget |
Sorbitol, % |
10,7 |
12,2 |
13,4 |
Sucrose, % |
1,79 |
2,70 |
3,60 |
Transporteret 14C (% af optaget) |
44,0 |
53,6 |
59,6 |
På lignende vis har velbelyste træer ved et givet frugt/blad-forhold tykkere blade og større og mere tørstofrige frugter end skyggede planter (bilag 6-11). Dette svarer til de postulerede sammenhænge i afsnit 6.2.2. Under markforhold er lys en meget vigtig faktor, som påvirker source-aktiviteten. Lysforholdene påvirkes bl.a. via plantesystemer og beskæring.
Bilag 6-11. Virkning af lysniveau på blad- og frugtudvikling. 'Golden Delicious' (Hansen 1977).
Vækstforhold |
Friland |
Under plast |
Temperatur, gns. dag |
17,0 |
20,7 |
Antal frugter/hg blade |
53 |
48 |
Bladtykkelse, cg/dm² |
84 |
57 |
Frugtstørrelse, g/frugt |
100 |
78 |
Farve (100 = helt gul) |
53 |
35 |
Total tørstof, % |
11,9 |
9,7 |
Opløselig tørstof, % |
8,3 |
5,3 |
Skold, % |
26 |
7 |
Som allerede nævnt indgår kalium i relativt store mængder i frugter. Frugter, som er velforsynede med kalium, vil derfor vokse hurtigere end frugter hos træer, som mangler kalium (bilag 6-12). Samtidig vil de velforsynede frugter hurtigere kunne omsætte assimilater. Dette fører til et større "træk" i disse, og det virker fortyndende på systemet.
Bilag 6-12. Virkning af kaliumforsyning på frugtstørrelse og tørstofindhold (Hansen 1982).
Antal frugter pr. træ |
g pr. frugt |
% opløseligt tørstof ved: |
|||
Kaliumtilførsel |
80 g |
130 g |
180 g |
||
Normal |
37 |
117 |
15,1 |
14,7 |
14,7 |
Ingen |
35 |
98 |
15,9 |
16,3 |
16,4 |
Samtidig med større frugter ved god kaliumforsyning får vi altså et lavere tørstofindhold, når der sammenlignes med kaliummanglende frugter. Dette kan opfattes som et eksempel på virkning via frugtens sink-aktivitet. Frugtens sink-aktivitet påvirkes nok ofte af stoffer, som kommer fra roden, og som er af betydning også for frugtens vækst. Derfor vil virkninger på rod/top-forhold (beskæring) eller på rodstørrelse eller rodaktivitet ofte føre til virkninger på frugtens sink-aktivitet. Det vises der eksempler på i de senere kapitler (afsnittene 14.2, 14.8, 15.2, 15.3). Vækststoftilførsel vil også undertiden kunne påvirke frugternes sink-aktivitet, f.eks synes frugter med cytokininaktivitet at kunne øge frugtvæksten og nedsætte sukkerindholdet (Tartarini et al 1993).
Der er en betydelig sortsvariation, hvad frugtvæksthastighed angår. Hos 'Graasten' vil den således ved et givet frugt/blad-forhold ofte kunne være ca. 25% større pr. frugt end hos 'Golden Delicious'. Det skyldes tilsyneladende genetiske forskelle i frugternes sink-aktivitet, idet forsøg har udelukket forskellig bladaktivitet som årsag (Hansen 1977b).
Temperaturen påvirker inden for visse grænser al biologisk aktivitet, altså også frugtvækst. Det kan derfor hævdes, at også temperaturen er en sink-aktivitetsfaktor. Temperaturens hovedvirkning ligger dog måske i at påvirke det tidsmæssige udviklings- og modningsforløb (afsnit 7.3.1).
6.6. Vekselvirkning mellem source-sink-faktorer (æble)
Der gøres forsøg på at udtrykke frugtvækstsammenhængen matematisk. Udbyttet pr. ha (y) som funktion af frugtantal pr. ha (x), som illustreret i bilag 6-9, kan muligvis udtrykkes som
y = kx / K+x
svarende til Michaelis-Menton-relationen i enzymkinetik. Her kan man muligvis opfatte k som et udtryk for source-aktiviteten og 1/K som et udtryk for sink-aktiviteten. (Det må dog bemærkes, at på hektarbasis vil en ændring af løvdækket også kunne påvirke k og K, men med samme faktor.)
Hvis ovennævnte ligning gælder, kan det vises, at en lignende sammenhæng vil gælde for relationen mellem frugtstørrelse og blad/frugt-forhold (bilag 6-8), og at frugtstørrelsen som funktion af udbytte pr. ha vil danne en ret linje, hvor hældningskoefficienten angiver -1/K, skæringspunktet med abscissen angiver k, og skæringspunktet med ordinaten svarer til k/K (= potentiel frugtstørrelse). Bilag 6-13 viser, hvordan relationen mellem frugtstørrelse og udbytte kan tænkes ændret med plantagens alder, hvis der ikke sker større indgreb ved beskæring og lignende. Sammenhængen er særlig interessant, fordi udbyttet er en direkte økonomisk parameter, og det samme gælder frugtstørrelsen via dens sammenhæng med sorteringsudfaldet.

Bilag 6-13. Sammenhænge mellem frugtstørrelse og udbytte. Postulerede ændringer ved ændring i alder (på grund af øget løvdækning), samt hvis der i de ældre samtidigt forudsættes dårlige lysforhold (reduceret source-aktivitet) samt ydermere dårlig vækst (reduceret sink-aktivitet).
I en østtysk undersøgelse (Schuricht 1986) med almindelige dyrkningsforhold blev det over en 18-årig periode vist, at frugtmængden var den enkeltfaktor, som kraftigst påvirkede frugtstørrelsen. Der var også en positiv korrelation med sommernedbøren (virkning via sink-aktivitet), såvel som med temperaturen (virkning ved at påvirke frugtudviklingsperiodens længde (afsnit 21.2) eller ved at øge frugtudviklingshastigheden (afsnit 7.3.1)).
6.7. Blomsterkvalitet, fysiologisk kvalitet af unge frugter
Frugtsætningen kan afhænge af blomsterkvaliteten, som igen afhænger af forskellige faktorers indvirkning på blomsterudvikling, oplagsnæring mv. som omtalt i tidligere kapitler. Måske kan blomsterkvaliteten også have betydning for den senere frugttilvækst, selv om det nok er en mindre vigtig faktor, når hele træer sammenlignes. Ved vekselbæring kan de unge frugter dog have et lavere celleantal året efter et bæreår.
Frøantallet afhænger af bestøvningseffekten, men frugttilvæksten afhænger ikke eller kun i ringe omfang af frøantallet hos æble (modsat jordbær, Ribes m.fl.).
6.8. Frugtstørrelsesvariation inden for et træ (æble)
I de foregående afsnit er det vist, hvordan virkninger på træet som helhed via source-sink-relationerne kan påvirke frugters vækst og indhold. Men også inden for et træ er der oftest en betydelig variation, hvad frugtstørrelse, dvs. frugtvækst, angår.
Variationen i koncentration af sukker, syre mv. er langt mindre. I lyset af de foranstående afsnit ville lokale forskelle i blad/frugt-forhold kunne påvirke frugtvæksten. Det ser dog ikke ud til at være en væsentlig faktor, i det mindste ikke i mindre træer (bilag 6-14, se også bilag 5-2e og g).
Bilag 6-14. Frugtudvikling ved lokale forskelle i blad/frugt-forhold (Hansen 1982).
Antal frugter pr. spore |
g pr. frugt |
% total tørstof |
% opløs. tørstof |
% titr. syre |
1 |
61,5 |
15,4 |
10,8 |
1,12 |
2 |
63,8 |
15,1 |
10,8 |
1,10 |
3 |
64,1 |
15,2 |
10,8 |
1,07 |
4 |
63,7 |
15,3 |
10,9 |
1,09 |
I større træer kan variation i lysintensitet være årsag til betydelige forskelle i frugtudvikling. I de ydre dele øges bladenes fotosynteseintensitet og stivelsesindhold (bilag 6-15), og frugterne bliver større med et øget tørstofindhold (bilag 6-16). Derimod kan syreindhold og frugtens fasthed være mindsket; det kan skyldes en samtidig virkning på modningshastigheden (kap. 7). I mindre, åbne træer er der dog ikke tydelige forskelle i frugtudviklingen på forskellige grene.

Bilag 6-15. Fotosyntesehastighed og bladtykkelse i de ydre og indre kronedele af æbletræ. Efter Marini & Barden (1981).
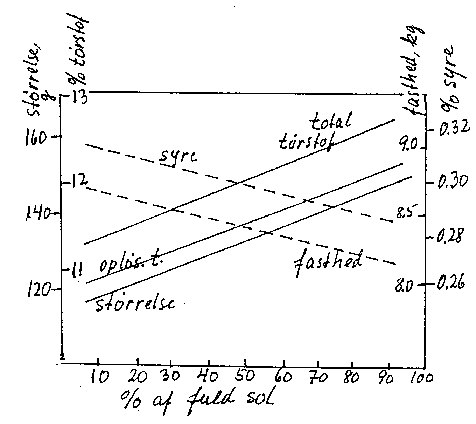
Bilag 6-16. Frugtudvikling hos æble som funktion af solindstrålingen i forskellige grenpartier. Efter Robinson et al. (1983).
Men alligevel er der også her en betydelig variation i frugtstørrelse inden for et træ. Der er en vis sammenhæng mellem størrelsen af unge frugter og den endelige frugtstørrelse. Forskelle i unge frugters fysiologiske kvalitet kan være knyttet til forskelle i blomsterkvalitet, som igen kan afhænge af skudtype. Fx kan frugter udviklet på sidste års årsskud være mindre end øvrige frugter (se bilag 2-15). Undertiden bliver frugter mindre (med lavere tørstof- og højere syreindhold) med stigende sporealder.
Men også ud over denne faktor kan der være en betydelig variation i frugtstørrelsen inden for et træ. Tilsyneladende må dette da skyldes forskelle i de enkelte frugters sink-aktivitet, dvs. forskelle i tilførsel af sink-faktorer. Muligvis kan den primigene dominans spille ind (afsnit 4.2.2.).
Forskelle i frugternes væksthastighed inden for et træ har betydning for virkningen af frugtudtynding.
6.9. Frugtvækst og -kvalitet hos blommer
Forholdene hos pære er næsten som hos æble, og også hos andre relativt storfrugtede arter (fersken, kiwi) er der tydelige sammenhænge mellem frugt/blad-forhold og frugtudvikling. Dette gælder også hos blommer, hvor mange udtyndingsforsøg har vist, at et reduceret frugtantal giver større frugter. Samtidig øges også indholdet af tørstof og sukker som vist i norske undersøgelser (bilag 6-17), der også har vist positive sammenhænge mellem frugtstørrelse, sukkerindhold og farveudvikling. Som det vil blive vist kan beskæring øge sink-aktiviteten (bilag 14-35), så der fås større frugter, men med et lavere tørstofindhold, og der kan være vekselvirkninger med bæringsgraden. Disse resultater tyder på, at der også hos blommer er mange lighedspunkter med frugtudviklingen hos æbler.
Bilag 6-17. Virkning af frugtantal på sukkerindhold (%) hos blommer (Vangdal 1982).
Antal frugter pr. blomsterstand |
Fructose |
Glucose |
Sucrose |
Sorbitol |
Total sukker |
0,1 |
0,8 |
2,2 |
7,6 |
0,3 |
11,1 |
0,4 |
0,6 |
2,1 |
5,1 |
0,2 |
8,2 |
0,6 |
0,6 |
2,0 |
3,8 |
0,2 |
6,8 |
1,0 |
0,6 |
2,1 |
2,4 |
0,2 |
5,6 |
6.10. Frugtvækst og frugtkvalitet hos kirsebær
Hos småfrugtede arter har de enkelte frugter en genetisk betinget mindre sink-aktivitet (vækstaktivitet). Derfor skal der mange frugter til at give en samlet set stor sink-styrke. Derfor er virkningen af frugtantal (frugt/blad-forhold) på frugtvækst og -kvalitet langt mindre end hos storfrugtede arter. Hos den meget småfrugtede surkirsebærsort 'Stevnsbær' giver selv en stor reduktion af bærantallet ingen eller kun en lille forøgelse af bærstørrelsen (bilag 6-18), og der er ingen tydelige virkninger på bærrenes sammensætning. Hos mere storfrugtede surkirsebærsorter og hos sødkirsebær er der dog ved et reduceret frugtantal fundet lidt større frugter med højere tørstof- og farveindhold. Source-aktiviteten kan også spille ind, skygning kan give langsommere frugttilvækst og frugter med lavere tørstof- og farveindhold. Ved beskæring (se afsnit 14.8.2) øges sink-aktiviteten med større, men mindre tørstofrige frugter til følge.
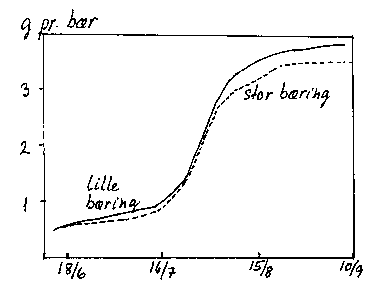
Bilag 6-18. Frugttilvækst hos surkirsebær ('Stevnsbær) ved lille og stor bæring (hhv. 7,0 og 24,9 kg bær pr. træ). Pometet.
Ved sammenligning inden for et træ giver de tidligst udviklede blomster større frugter med et højere tørstof- og farveindhold. Dette tyder på, at blomsterkvaliteten også kan have betydning for frugtudviklingen.
6.11. Frugtvækst og frugtkvalitet hos solbær (Ribes)
Solbær har endnu mindre frugter end kirsebær. Alligevel giver et mindsket frugt/blad-forhold større frugter (bilag 6-19). Dette skyldes nok, at frugterne her sidder sammen i klaser, som får tilførsel samlet, så at der lettere opstår konkurrence.

Bilag 6-19. Frugttilvækst ved forskellige behandlinger af solbærbuske (Hansen & Ostermann 1988).
Det er dog næppe først og fremmest via en direkte virkning på assimilattilgængeligheden, at frugtvæksten er påvirket, da større frugter efter en frugtudtynding følges af et reduceret tørstofindhold (bilag 6-20). Måske fremmer den større assimilatrigelighed efter en frugtudtynding generelt tilvæksten, herunder rodaktivitet, så tilførslen af sink-faktorer til frugten øges. Frugttilvæksten påvirkes nemlig tydeligt ved indvirkninger på top/rod-forhold og rodaktivitet, som påvirker sink-aktiviteten (bilag 6-19), og der er en positiv sammenhæng mellem bærstørrelse og skudvækstintensitet (bilag 6-21).

Bilag 6-20. Sammenhæng mellem bærstørrelse og total tørstof-, hhv. syreindhold hos solbær (under forskellige vækstpåvirkninger) (Hansen & Ostermann 1988).
Frugtstørrelse og koncentration af totalt og opløseligt tørstof er negativt korreleret (bilag 6-20), som det postuleres ved virkningen via sink-aktiviteten. Dog stiger syreindholdet parallelt med frugtstørrelsen. Ved meget tæt plantning bliver bærstørrelsen mindre (bilag 4-13). Dette kan også skyldes lysmangel (virkning vis source-aktivitet). Ligeledes er tørstofindholdet lavere i bærrene i den indre del af busken.
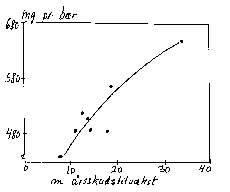
Bilag 6-21. Sammenhæng mellem bær- og skudtilvækst hos solbær (forskellige vækstpåvirkninger) (Hansen & Ostermann 1988).
Virkninger på buskens tæthed kan muligvis også virke lidt ind på bærrenes tørstofprocent ved at påvirke bærrenes transpirationsintensitet.
Ved sammenligning af buske er sink-aktiviteten altså den væsentligste faktor for virkninger på frugtvækst. Den påvirker således svulmningen af bærrene, store frugter har den største frugtmængde pr. frø. Inden for planten, og det vil især sige inden for klasen, er blomsterkvaliteten den væsentligste faktor med hensyn til bærvækst. Frøantallet har nemlig betydning for bærvæksten; trods betydelig variation stiger den maksimalt opnåelige bærstørrelse med frøantallet (bilag 6-22). Frøantallet er især afhængigt af antallet af æg, som bestemmes af blomsterudvikling og blomsterkvalitet. Bestøvningseffekten spiller dog også ind. De inderste bær på klasen bliver som helhed større end de yderste (bilag 6-22), men det skyldes for en stor del mere veludviklede blomster med flere æg, dog vil de også stå stærkere i konkurrencen inden for klasen under bærrenes vækst (sammenlign blomster- og bærfald i afsnit 4.3.3).

Bilag 6-22. Sammenhæng mellem bærvægt og frøantal, henholdsvis bærrets position. Linjerne viser det potentielle niveau. Efter Webb (1972)
.Foruden forskelle i blomsterkvalitet må der også være en betydelig variation mellem de enkelte frugters sink-aktivitet, da der ved et givet frøantal forekommer bær af forskellig størrelse, dvs. med forskellig svulmingsgrad for frø (bilag 6-22).
6.12. Frugtvækst og frugtkvalitet hos jordbær
Mens blomsterkvaliteten hos solbær spiller en stor rolle for bærvæksten, især for størrelsesvariationen inden for en klase, så er blomsterudvikling og blomsterkvalitet afgørende for bærstørrelsen hos jordbær, som altså for en stor del er bestemt allerede ved blomstringen.
6.12.1. Blomsterkvalitet (antal æganlæg pr. blomst)
Følgende relation forklarer størstedelen af variationen i bærstørrelse under givne omstændigheder:
Bærvægt (g) = (total achener/bær - C) x F / antal achener/cm² (Abbott, Webb et al.).
Bærvægten er derfor en retlinet funktion af acheneantal/bær ved en given achenetæthed (bilag 6-23) eller bærvægt x achenetæthed er en retlinet funktion af acheneantal/bær. C angiver en numerisk korrektion på grund af, at bær ikke er sfæriske, den varierer med sorter, men også kan påvirkes af dyrkningsbetingelser.
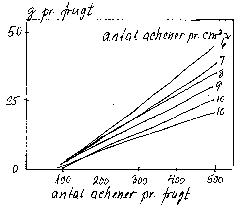
Bilag 6-23. Bærvægt hos jordbær som funktion af acheneantal og achenetæthed (Abbott et al. 1977; Webb et al. 1978).
F har dimensionen g/cm² og siger i virkeligheden, at et jordbær kan opfattes som sammenfoldet væv af en given tykkelse (F), som afhænger af sorten, men som også kan påvirkes af dyrkningsforhold. Ved større bær stiger andelen af luftrum i bærret, i store bær kan der optræde egentlige hulrum.
Bærstørrelsen afhænger derfor af antallet af æganlæg (støvveje), som sidder på den hvælvede blomsterbund, som senere danner frugtkødet. Blomsterudviklingen (se afsnit 2.6.3) bestemmer antallet af æganlæg pr. blomst; den er bl.a. genetisk bestemt.
Ydre faktorer (dyrkningsfaktorer) påvirker nok især blomsterudviklingen via virkningen på konkurrenceforholdene mellem eller inden for planten. Aftagende planteafstand ved sommerplantning kan give mindre bær næste år (trods færre blade og blomsterstande pr. plante, bilag 13-19). Kraftig kvælstoftilførsel giver flere blade og blomsterstande pr. plante, men samtidig mindre bær (afsnit 19.2.1). Ældre planter får flere kroner pr. plante og mindre bær. Det ser ud til at når antallet af blomsteranlæg pr. plante stiger, evt. også når antallet af blade øges, så stiger den interne konkurrence om stoffer eller faktorer, som virker ind på blomsterudviklingen, så at der fås blomster af en dårligere kvalitet (færre æganlæg pr. blomst). Dette fører til, at der kan påvises tydelige negative sammenhænge mellem bærantal pr. plante og bærstørrelsen, f.eks. i 'Elsanta' (Rindom & Hansen 1995).
Inden for planten får de tidligst initierede blomsterstande flest blomster, men alligevel er bær af en given orden undertiden alligevel større end hos senere initierede stande. Bærstørrelsen er i meget høj grad knyttet til blomsterorden (bilagene 2-20, 6-24), men denne variation kan stort set forklares ved en parallel variation i æganlæg pr. blomst (afsnit 2.6.3). Man har været inde på, at summen af bærvægten eller acheneantal pr. bær sammenlagt pr. blomsterorden er konstant, dvs. de skulle pr. bær aftage med blomsterorden i forholdet 1:1/2:1/4:1/8 . . . for primære, sekundære bær etc. Så konsekvent er sammenhængen dog ikke; genetiske og andre forhold kan spille ind.

Bilag 6-24. Bærtilvækst hos jordbær i afhængighed af blomsterorden.
Fasciation, hvor flere bær udvikles i sammenvokset stand, er knyttet til uheldige indvirkninger på blomsterudviklingen efteråret i forvejen. Der kan også være dele af blomsten, som ikke udvikles ordentligt; virkninger på plantematerialet, som præger det i mere vegetativ retning, kan forøge andelen af misdannede bær.
6.12.2. Forholdet achener/æganlæg
Bestøvning og befrugtning har virkning på dette forhold og dermed på bærstørrelsen. Fjernes støvvejene på dele af blomsterbunden, udvikles den ikke i dette område; bærrene bliver skæve. Tilføres der auxin, genetableres væksten (bilag 6-25). Udvikling af achener (frø) har altså betydning ved, at de afgiver vækststoffer, som er nødvendige for bærrets vækst. Mangelfuld bestøvning (afsnit 3.2.3) eller frostskade, hvor blot nogle af griflerne ødelægges, er nok hovedårsagerne til misformede bær.

Bilag 6.25. Bærvækst hos jordbær som funktion af acheneantal og auxin-tilførsel. A) normal; B) frø fjernet fra siderne; C) alle frø fjernet; D) frø fjernet + auxin-tilførsel.
På et bær kan der ofte ses achener af forskellig størrelse (udviklingsgrad), uden at det tilsyneladende har influeret på bærrets vækst. Måske er det ikke i alle tilfælde nødvendigt med fuld acheneudvikling, for at den maksimale virkning på bærvækst kan ske.
6.12.3. Bærvækst, bærsvulmning pr. achene
Frøantallet pr. frugt er som nævnt en vigtig faktor for frugtvækst, men også faktorer under selve bærvæksten spiller ind, og dette resulterer i forskellig achenetæthed (bilag 6-23). Maksimal bærsvulmning giver achenetæthed på 6 achener/cm², men værdier på 8-12 er almindelige, så svulmningen er ofte under den maksimale.
Ved sammenligning af forskellige dyrkningsforhold vil følgende faktorer kunne spille ind.
Blad/frugt-forhold synes ikke at være en afgørende faktor. Fordi udtynding af frugter giver ingen eller relativ lille forøgelse af bærtilvæksten (bilag 6-26). Lidt lavere achenetæthed og lidt større tørstofprocent tyder dog på en svag positiv virkning af større assimilattilgængelighed, denne har også resulteret i en større bladtilvækst.
Bilag 6-26. Frugtudvikling og bladtilvækst efter fjernelse af ca. halvdelen af blomsterne hos jordbær (repræsentativt fordelt på forskellig blomsterorden).
Ubehandlet |
Udtyndet |
|
Bærstørrelse, gns. g/frugt |
9,01 |
9,06 |
Achenetæthed, antal/cm² |
10,0 |
9,5 |
Total tørstof, % |
10,6 |
11,1 |
Bladmasse, g tørvægt/plante |
37 |
44 |
Source-aktivitet. Bladenes gensidige skyggevirkninger er nok oftest begrænsede, bladaktiviteten hører næppe til de særligt begrænsende faktorer. Reduktion af lysintensiteten til ca. det halve af normal indstråling, såvel som fjernelse af ca. halvdelen af bladene, påvirkede ikke bærstørrelsen i 'Elsanta', når behandlingerne indskrænkede sig til 3-4 uger under selve bærudviklingen, derimod øgedes bærstørrelsen med ca. 10%, hvis halvdelen af blomsterstandene eller af kronerne blev fjernet.
Sink-aktivitet er nok den faktor, som især har betydning for bærsvulmning og achenetæthed. Konkurrencen mellem stande, eller måske mellem blade og stande, fx om rodfaktorer kan reducere bærsvulmningen. Vandforholdene er måske den vigtigste faktor; tørkeperioder under bærudviklingen resulterer i mindre bær (bilag 20-23). Den aftagende bærstørrelse med plantealder skyldes måske også delvis en større achenetæthed.
Blomsterorden. Inden for blomsterstanden er det ikke blot sådan, at acheneantal og initial størrelse aftager med blomsterorden, men det gør også den absolutte såvel som den relative væksthastighed, inklusive hastigheden af overfladeforøgelse pr. achene (bilag 6-24, Forney & Breen 1985). Måske kan det være virkninger knyttet til blomsterkvaliteten, men det skyldes nok snarere konkurrencen mellem frugterne inden for standen, hvor primære og dernæst sekundære frugter dominerer eller har en fortrinsstilling, hvadenten det drejer sig om konkurrence om assimilater eller sink-faktorer. Fjernes de primære og sekundære frugter, kan det resultere i, at de øvrige frugter vokser hurtigere, mens det omvendte næppe er tilfældet.
Med jordbær er afsluttet en række eksempler, som viser, at vægten af forskellige frugtvækstfaktorer varierer med frugtarten (bilag 6-6), bl.a. under hensyn til genetisk bestemt frugtstørrelse, antallet af frugter i frugtstanden samt dennes opbygning.
6.12.4. Bærkvalitet
Smagsværdien hos jordbær afhænger især af aromastoffer (kap.8), sukker og syreindhold, som i høj grad er genetisk bestemt. Mens sukker- og syreindhold i f.ex. æble som allerede nævnt kan påvirkes meget af source/sink-forhold og de faktorer, som indvirker herpå, så er indvirkningen af fysiologiske, og dyrkningsmæssige faktorer på sukker- og syreindhold hos jordbær meget ringe. Tørstofindholdet øges ved vandmangel. Ved en given plukkedato kan de store frugter have et lidt højere sukkerindhold end de mindre bær, som har haft en mindre gunstig position og derfor måske lidt dårligere assimilatforsyning. De seneste bær under plukkeforløbet kan også vise lidt stigende sukkerindhold, måske fordi blad/frugt-forholdet så tiltager. Små planter giver undertiden frugter af lidt højere tørstofkoncentration end større planter. Men de nævnte virkninger er relativt små (Hansen 1995). Større planteafstand kan øge tørstofindholdet (afsnit 13.7.)
Westwood, M.N. 1978. Temperate-zone pomology. San Francisco. 428 pp.
Keipert, K. 1981. Beerenobst. Stuttgart. 349 pp.
Anon. 1984. Håndbog om frugt og grønsager. Bioteknisk Institut, Kolding.
Vestrheim, S. 1986. Sukker og nærslekta emne i frukt og bær. Frukt og bær. Oslo, pp. 56-66.
Roemer, K. 1989-92. Das Zuckermuster verschiedener Obstarten. Erw.Obstbau 31:211-216, 32:7-12, 42-46, 218-221, 33:169-172, 34:198-200.
Patrick, J.W. 1976. Hormone-directed transport of metabolites. I: J.F. Wardlaw & J.B. Passioura (eds.). Transport and transfer processes in plants, pp. 433-446.
Hansen, P. 1982. Assimilation and carbohydrate utilization in apple. Proc. 21st Int. Hort. Congr. Vol. I, pp. 257-268.
Hansen, P. 1977a. Blad-frugt relationer og frugtudvikling hos æbletræer. Tidsskr. Planteavl 81:123-132.
Johnson, D.S. 1992. The effect of flower and fruit thinning on the firmness of "Cox's Orange Pippin" apples at harvest and after storage. J.Hort.Sci. 67:95-101.
Hansen, P. 1977b. The relative importance of fruits and leaves for the cultivar-specific growth rate of apple fruits. J. Hort. Sci. 52:501-508.
Tartarini, S., S. Sansavini & M. Ventura 1993. CPPU control of fruit morphogenesis in apple. Scient.Hort. 53:273-279.
Schuricht, R. 1986. Die Wirkung von Ertrag, Witterung und Baumalter auf die Fruchtgrösse bei Apfel. Arch. Gartenbau 34:389-405.
Martini, R.P. & J.A. Barden 1981. Seasonal correlations of specific leaf weight to net photosynthesis and dark respiration of apple leaves. Photosynthetic Research 2:251-258.
Robinson, T.L., E.J. Seeley & B.H. Barritt 1983. Effect of light environment and spur age on 'Delicious' apple fruit size and quality. J. Amer. Soc. Hort. Sci. 108:855-861.
Vangdal, E. 1982. Sugars and sugar alcohols in Norwegian-grown plums. Meld. Norges Landbrulshøgskole 61, nr. 12. 39 pp.
Måge, F. 1994. Fruit development of Victoria plums in relation to leaf number. Acta Hort. 359:190-194.
Hansen, P. 1993. Source/sink relations in fruits. VI. Fruit/leaf-ratio and fruit decelopment in sour cherry "Stevnsbær". Gartenbauwiss. 58:134-136.
Roper, T.R. & W.H. Loescher 1987. Relationships between leaf area per fruit and fruit quality in 'Bing' sweet cherry. Hort. Sci. 22:1273-1276.
Patten, K.D. & E.L. Proebsting 1986. Effect of different artificial shading times and natural light intensities on the fruit quality of 'Bing' sweet cherry. J. Amer. Soc. Hort. Sci. 111:360-363.
Patten, K.D. & M.E. Patterson 1986. Factors accounting for the within-tree-variation of fruit quality in sweet-cherries. J. Amer. Soc. Hort. Sci. 111:356-360.
Hansen, P. & J. Ostermann 1988. Source-sink relations in fruits. II. Fruit growth and composition in black currants (Ribes nigrum L.). Gartenbauwiss. 53:155-159.
Webb, R.A. 1972. Fruit size in black currant and the assessment of crop potential. J. Hort. Sci. 47:199-204.
Abbott, A.J., G.R. Best & R.A. Webb 1977. The relation of achene number to berry weight in strawberry fruit. J. Hort. Sci. 45:215-222.
Webb, R.A., J.H. Terblanche, J.V. Purvis & M.G. Beech 1978. Size factors in strawberry fruit. Scient. Hort. 9:347-356.
Forney, C.F. & P.J. Breen 1985. Growth of strawberry fruit and sugar uptake of fruit discs at different inflorescence positions. Scient. Hort. 27:55-62.
Hansen, P. 1989. Source-sink relations in fruits. IV. Fruit number and fruit growth in strawberries. Acta Hort. 265:377-381.
Rindom, A. & P. Hansen 1995. Effects of fruit number and plants status on fruit size in strawberry. Acta Agric.Scand.
Hansen, P. 1995. Effects of flower and fruit development and cultural factors on fruit composition in strawberry. Acta Agric.Scand.