KAPITEL 4. Frugtsætning
Index
4.2.1. Hunlig sterilitet
4.2.2. Blomsterkvalitet i øvrigt (EPP)
4.2.3. Kirsebær
4.3.1. Æble
4.3.2. Kirsebær
4.3.3. Ribes
4.4. Frø- og frugtudvikling. Parthenocarpi. Vækststoffer
Når bestøvning og befrugtning er sket, begynder blomstens udvikling mod en frugt. Selve frugtsætningsprocessen kan deles op i trin, og forskellige faktorer kan virke ind på forløbet. Man taler dels om den initiale sætning, som måles ca. 4 uger efter begyndende blomstring, og som stort set er et udtryk for bestøvningseffekten, og dels om den endelige sætning. Forskellen mellem de to udgør det såkaldte junifald. Sætning måles i antal frugter pr. blomsterstand ved æble og pære, ellers som frugtantal i procent af blomsterantal.
Blomstermængden er naturligvis en afgørende forudsætning for den samlede endelige sætning. Der kan dog ske en vis kompensation ved en øget sætning pr. blomsterstand ved ringe blomstertæthed og under gode bestøvningsforhold især på grund af mindre junifald (storfrugtede arter).
Følgende faktorer kan spille en afgørende rolle i frugtsætningsforløbet:
Forenelighed mellem pollen og støvvej er et særdeles vigtigt punkt. Der findes selvfertile, delvis selvfertile (selvsterile), selvsterile og intersterile sorter (se kap. 3).
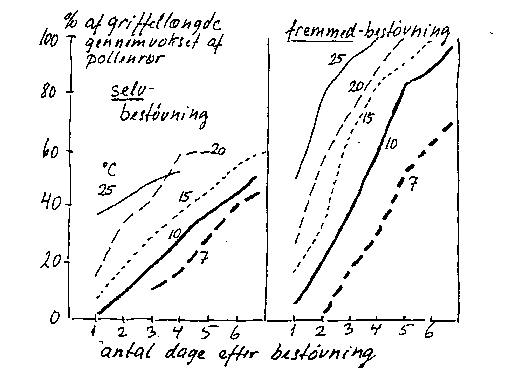
Dårlig forenelighed, fx med sortens eget pollen, viser sig ikke så meget under pollenkornenes spiring på støvfanget, men især ved at pollenrørenes vækst i griffelvævet går langsommere og eventuelt i stå i modsætning til foreneligt pollen (bilag 4-1). I enkelte tilfælde når dog eget pollen hos selvsterile sorter ned i frugtknuden.
Bilag 4-1. Pollenrørsvækst ved selv- og krydsbestøvning ved forskellige temperaturer i pære. Efter Vasilakakis & Porlingis (1985).
Selvsterilitet er nemlig ikke altid absolut (pseudocompatibilitet kan forekomme). Hos den selvsterile 'Cox's Orange' øges pollenrørsvæksten hos eget pollen ved stigende temperatur (bilag 4-2) og ved rigelig pollenafsætning, dvs. ved godt vejr under blomstringen. Som nævnt i afsnit 3.1.3 kan selv sorter som 'Cox's Orange' og 'Comice' undertiden give god sætning, selv om der ikke er bestøvere nærved (bilag 3-7). Bartlett (Williams)-pæren kræver ikke bestøvere under varme klimaforhold (Californien), men derimod under køligere forhold (Oregon).
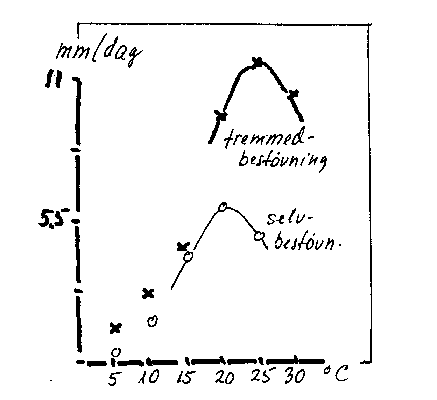
Bilag 4-2. Temperaturens virkning på pollenrørs væksthastighed efter selv- og fremmedbestøvning af 'Cox's Orange'. Efter Williams & Maier (1977).
Parthenocarpi (frugtdannelse uden frøudvikling) kan også i nogle tilfælde bidrage til frugtsætning ved manglende fremmed bestøvning. Ved afdækning af træer mod bier under blomstringen havde i gennemsnit af flere æblesorter og år 63% af dannede frugter ét til flere frø, mens 36% var parthenocarpe. Hos pærer var tallene 2 og 98%, da pærer lettere danner parthenocarpe frugter end æble (Karnatz 1963). Der var stor sorts- og årsvariation; i enkelte tilfælde var frugtantallet næsten lige så højt uden som ved fremmedbestøvning, i de fleste tilfælde dog betydeligt under.
En dansk undersøgelse i 'Summerred' viste også, at der i enkelte år, men langt fra i alle, kunne være et rimeligt udbytte uden fremmedbestøvning (bilag 4-3). Årsagen var dels parthenocarpe frugter, dels selvbestøvede frugter med frø, men med et langt lavere frøantal end ved fremmedbestøvning.
Bilag 4-3. Frugtsætning hos 'Summerred' uden og med fremmedbestøvning (Hansen 1989).
kg pr. træ |
% frøløse frugter |
Gns. frø/frugt i resten |
|
1980 Afdækket under blomstring |
9,0 |
51 |
1,2 |
----Fremmedbestøvning |
11,5 |
0 |
7,1 |
1985 Afdækket under blomstring |
5,1 |
58 |
1,3 |
----Fremmedbestøvning |
9,2 |
0 |
7,9 |
En del frugtarter er som tidligere nævnt normalt selvfertile. Dog kan man også i f.x. Ribes og Rubus sommetider observere, at krydsbefrugtning giver en bedre sætning end selvbestøvning. I en dansk undersøgelse i hyld blev sætningsprocenten således 68-77% ved fremmedbestøvning mod 35-52% ved selvbestøvning. Det er også vist, at i frugter med flere frø (ex. blåbær) kan frøantallet og frugtstørrelsen være større med fremmed- end ved selvbestøvning.
Spørgsmålet om grader af selvfertilitet kan muligvis inddeles som følger:
a. Normalt selvsterile sorter (æble, pære): Eget pollen når normalt ikke ned til frugtknuden undtagen ved særligt gode betingelser for pollenrørsvækst. Desuden kan parthenocarpi forekomme hos nogle sorter i et vist omfang.
b. Delvis selvfertile/selvsterile sorter (visse blommesorter): Det er nok også især i år med gunstige bestøvningsbetingelser, at disse sorter optræder som selvfertile.
c. Normalt selvfertile sorter (Ribes, Rubus): Eget pollen når normalt ned til frugtknuden, undtagen under ugunstige bestøvningsbetingelser, idet der i nogle tilfælde kan være øget udbytte ved en fremmedbestøvning.
Ydre såvel som udviklingsmæssige forhold kan påvirke blomsterkvaliteten, dvs. frugtsætningsevnen.
4.2.1. Hunlig sterilitet
"Sterile" blomster kan forekomme, fx ved at nattefrostskade har ødelagt grifler eller andre dele (se kap. 24). En kold periode uden egentlig frost kan muligvis også skade. I f.ex. kirsebær kan en stor del af blomsterne slet ikke udvikle frugter (se senere).
4.2.2. Blomsterkvalitet i øvrigt (EPP)
Kernefrugt. I afsnit 2.6 er det beskrevet, hvordan blomsterudviklingen kan føre til blomster af forskellig kvalitet. Æganlæggene kan undergå en mere eller mindre fuldstændig udvikling. Det kan have betydning for deres levetid, som kan have forskellig varighed i antal dage, efter at blomsten er åbnet. Dette får betydning for EPP, den effektive pollinerings (bestøvnings)-periode, idet det tager en vis tid for pollenrørene at vokse ned (bilag 4-4). Jo længere EPP'er, desto større er chancen for en effektiv bestøvning og dermed en større frugtsætning.
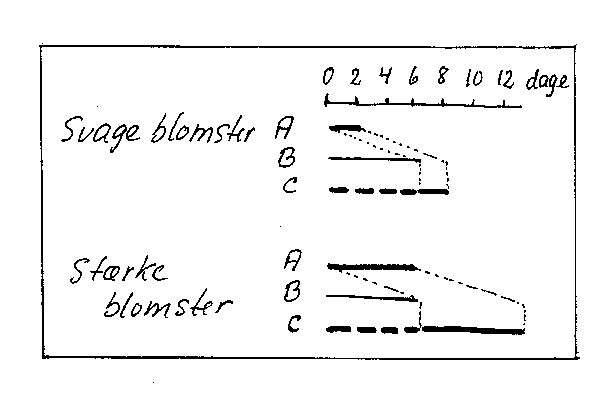
Bilag 4-4. Eksempler på kort og lang EPP (effektiv pollineringsperiode (A)) beregnet som difference mellem æganlægs levetid (C) og pollenrørs væksttid efter blomstens åbning (B). Efter Williams & Wilson (1970).
EPP kan være reduceret ved:
a. Stor bæring året før.
b. Kraftig skudvækst året før (bilag 4-5). Eventuelle blomster på årsskud har ofte en dårligere blomsterkvalitet (bilag 2-15). Ungt ved og helt unge træer har en dårligere frugtsætning og en større andel degenererede æganlæg end ældre ved og træer. Det samme gælder oprette i forhold til vandrette skud.
Bilag 4-5. Sammenhæng mellem skudvækst og frugtsætning.
Skudvækst: |
Svag |
Moderat- kraftig |
||
Bestøvergrene: |
-- |
+ |
-- |
+ |
1977 Skudvækstkarakter |
0 |
3 |
33 |
32 |
Blomstertæthed* |
95 |
92 |
81 |
82 |
Initial sætning** |
1,40 |
1,32 |
0,20 |
0,76 |
Endelig sætning** |
0,84 |
0,88 |
0,12 |
0,60 |
1978 Skudvækstkarakter |
23 |
-- |
67 |
57 |
Blomstertæthed* |
82 |
-- |
36 |
27 |
Initial sætning** |
1,42 |
-- |
0,75 |
1,83 |
Endelig sætning** |
0,93 |
-- |
0,70 |
1,23 |
* 100 = max. ** Antal frugter pr. blomsterklase.
c. Dårlige lysforhold (tætte træer) året før.
d. Regulær kvælstofmangel under blomsterudviklingen (se afsnit 19.2.1).
e. Tynde knopper og skud samt blomster af sideposition
kan have en ringere frugtsætningsevne (bilag 4-6)..
Bilag 4-6. Sammenhæng mellem spore- og knopstørrelse, og blomsterknopdannelse og frugtsætning. Æble.
f. Et lunt forår, som muligvis tærer på oplagsnæringen og giver dårligere ægudvikling, kan give en nedsat frugtsætning (Jackson & Hamer 1980).
g. Der kan være betydelige sortsforskelle. Pæresorten 'Comice' kan have en særlig lav EPP. Beskæring om vinteren, så at alle årsskud fjernes (og konkurrencen om oplagsnæringen dermed mindskes), kan her øge frugtsætningen (bilag 14-36).
Temperaturen behøver ikke at have nogen større indflydelse på EPP. Ved faldende temperatur falder pollenrørenes væksthastighed og æganlæggenes henfaldshastighed nemlig samtidig. Ved meget lave temperaturer når pollenrørene dog næppe ned til æganlæggene, inden disse degenererer.
Bemærk, at blomsterkvaliteten, og dermed EPP, i store træk påvirkes af forskellige faktorer i samme retning som selve blomsterknopdannelsen.
De primære blade (forårsblade) er allerede anlagt inde i knoppen. Udvikling af dem er nødvendig, for at der kan ske en normal frugtsætning (bilag 4-7). Fjernelse af bægerblade kan også reducere frugtsætningen (se afsnit 5.2).
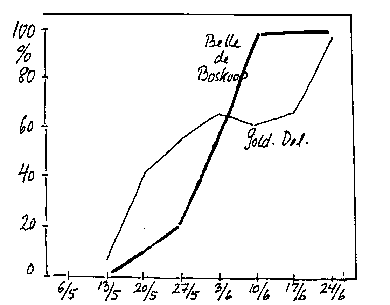
Bilag 4-7. Frugtsætning efter fjernelse af forårsblade på forskellige tidspunkter. Pct. af ubehandlet. Efter Dalbro (1966).
4.2.3. Kirsebær.
Også hos kirsebær kan blomsterne have forskellig kvalitet og frugtsætningsevne. Også her har de længste skud, hvor også blomsterknopdannelsen er dårligst, en ringere frugtsætningsevne (bilag 2-16). Det samme gælder knopper, som sidder i skygge (bilag 3-10).
Selv ved optimal bestøvning (håndbestøvning) sætter højst 50-60% af surkirsebærblomsterne frugt (bilag 3-10). Der kan være stor variation, og næsten sterile træer kan forekomme.
Undersøgelser på 'Stevnsbær' har vist, at manglende pollenrørsvækst kan være en medvirkende årsag til dårlig frugtsætning (bilag 4-8). Men en endnu større del af årsagerne er uopklarede. Måske skyldes det også her æganlæggenes levetid, så at de i nogle tilfælde går til grunde, inden pollenrøret når det sidste stykke gennem perikarpen (dette tager relativt lang tid). Polske undersøgelser tyder på, at pollenrørene i nogle tilfælde vokser ned i nærheden af æganlæggene, uden at det fører til befrugtning (Lech & Tylus 1983).
Bilag 4-8. Årsager til mangelfuld frugtsætning i 'Stevnsbær'. Efter Kühn (1988).
Teoretisk (alle befrugtede): |
100% |
Dårlig pollenoverførsel |
1-5 % |
Manglende pollenspiring |
1-10% |
Manglende pollenrørsvækst |
15-42% |
Andet (henfald af æganlæg) |
29-57% |
Befrugtning (reelt) |
16-27% |
Muligvis er der tale om forskellige grader, fra 1) reduceret blomsterknopdannelse, over 2) blomsteranlæg, som dør i knoppen (se bilag 2-18) og 3) tilsyneladende fuldt udviklede, men sterile blomster til 4) normale blomster, som kan befrugtes.
Stenfrugtblomster har oprindelig to æganlæg, hvor "normalt" det ene går til grunde, mens det andet udvikles. Men hvis dette anlæg også går til grunde, er der intet tilbage. Modsat æble, hvor der kan være op til 10 frøanlæg, så der stadig kan ske frø- og frugtudvikling, selv om halvdelen af æganlæggene skulle være defekte. Ved optimal bestøvning af æbleblomster kan der da også let opnås en initial sætning på over 90% af blomsterne.
Junifald forekommer især i storfrugtede arter og skyldes formentlig konkurrence om assimilater eller andre stoffer. De i forvejen dårligt stillede frugter (fx de mindste) vil have en mindre tiltrækningsevne. Dette fører til mindre vækst (evt. også mindre vækststofproduktion) hos disse, så at forskellen til de gunstigt stillede frugter øges, til sidst falder de ugunstigt stillede af. Måske er der tale om "primigenic" dominans, de mest udviklede frugter har en større polær transport af auxin (IAA), som på forbindelsesstederne til mindre udviklede frugter modvirker deres polære auxin transport. Dette bevirker så, at disse frugter får et dårligere udviklet ledningsvæv, derfor ringere transport af assimilater m.v. ind i frugten, og endelig vil en ringe polær auxin tranport fra frugten selv føre til dannelse af løsningslag i dens stilk (Bangerth 1989).
Dårlig xylemudvikling i frugtstilken er påvist i frugter som falder af.
Junifald øges derfor af:
a) En stor samlet initialsætning. Ved tiltagende initial sætning på træet som helhed (gennemsnit af alle sporer) nærmer den endelige sætning sig en "grænseværdi", et udtryk for, at træet kun kan understøtte en vis frugtmængde (bilag 4-9).
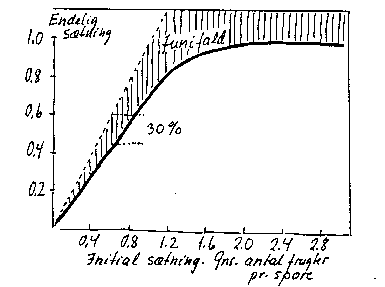
Bilag 4-9. Endelig sætning som funktion af initial sætning, begge som gennemsnit af samtlige sporer på træet. Æble. Efter Hansen (1980).
b) Ved stor skudvækstaktivitet. Sætningen kan øges, hvis skudspidserne fjernes (bilag 4-10).
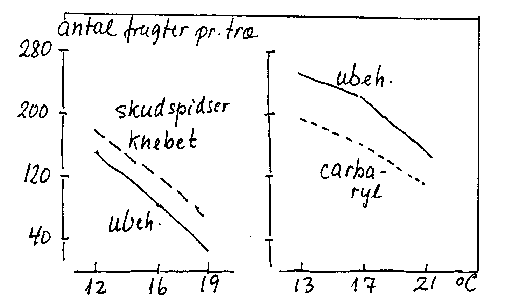
Bilag 4-10. Virkning af tidlig skudknibning, carbarylsprøjtning og temperatur på frugtsætningen hos æble. Efter Grauslund (1968).
c) Ved dårlige lysforhold i træet (lavt substratniveau). Skygning eller tilførsel af fotosynteseinhibitorer øger frugtfaldet.
d) Ved øget temperatur (uden tilsvarende lyskompensation) i perioden inden junifaldet (bilag 4-10). Nok fordi den samtidigt øgede vækstaktivitet forstærker konkurrencesituationen.
e) Ved vandstress (se afsnit 20.2.4).
f) Ved behandling med udtyndingsmidler (bilag 4-10 samt afsnit 15.5).
Foruden konkurrenceforholdene i træet kan blomsterudvikling, og dermed blomsterkvalitet, bestøvninges effektivitet og lignende, også spille ind.
4.3.2. Kirsebær
Junifald ("Rötel") kan forekomme ved meget stor initial sætning, men i mange tilfælde er junifaldet lille (bilag 3-10). I enkelte år kan der dog være et ekstraordinært frugtfald, som så tilsyneladende har forbindelse med de klimatiske forhold. Det kunne være knyttet til en unormal frøhvideudvikling, fx at frøhviden bliver diploid i stedet for som normalt triploid, men det synes ikke at være hele årsagen (Schanz & Stösser 1989).
Hos solbær kan der ske en stor abort af frø i de enkelte frugter (op til 40% af frøene eller undertiden mere).
Drysning, dvs. blomster- eller bærfald på 10-50% inden for de første 3-4 uger efter blomstringen kan være et alvorligt problem i solbær (Daugaard 1981, bilag 4-11). I ribs er det vist, at faldet er størst yderst på klaserne hvor blomsterne også har den dårligste kvalitet. Bestøvningen kan virke ind, men fænomenet kan forekomme også ved god bestøvning. Blomsterne yderst på klasen er dårligst udviklede og har det laveste antal frøanlæg, og disse blomster/bær er mest udsat for at falde af.
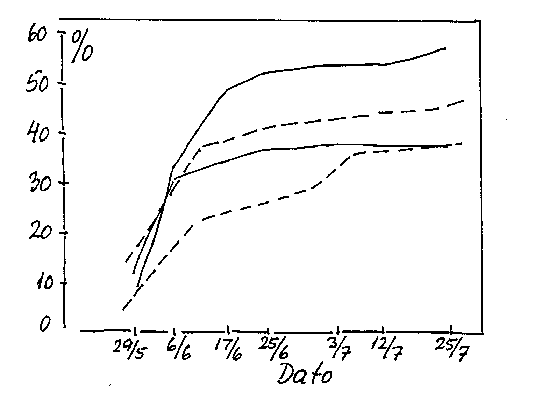
Bilag 4-11. Frugtfald (pct. af blomsterantal) hos solbær; resultater fra forskellige sorter og vækstforhold i 1985 og 1986. Efter Kühn (1988).
Selv om Ribes har relativt små frugter, så synes det forhold, at der sidder mange frugter i en blomsterstand at bevirke, at der opstår konkurrence inden for standen. De mindst begunstigede frugter sakker bagud, har eller får måske mindre vækststofproduktion, og falder derfor af. Hvis nærtsiddende blade fjernes, øges faldet, mens fjernelse af konkurerende basisskud mindsker faldet. Dette tyder på, at tilførsel af eller konkurrence om assimilater spiller ind. Mekanismen synes således at minde om den mekanisme, som gør sig gældende for junifald i æble, så måske vil der altid være et vist naturligt fald.
Faktorer, som påvirker konkurrenceforholdene i klasen (planten), skulle derfor påvirke frugtfaldet. I danske forsøg med vanding og tilførsel af vækstregulerende stoffer lykkes det ikke at reducere frugtfaldet (Kühn 1988). Ved meget kraftig væksthæmning synes frugtfaldet dog at være reduceret (bilag 4-12). I tætte buske med lysmangel 15. november 1996 reduceres frugtsætningen ligesom blomsterknopdannelse og frugtstørrelse (bilag 4-15). (Måske kan bestøvningsmulighederne også være dårligere under sådanne betingelser.)

Bilag 4-12. Virkning af arring (meget kraftig vækstreduktion) og etephon-sprøjtning på bærantal pr. klase hos solbær, sammenlignet med "almindelig" kultur. Efter Hansen & Ostermann (1988).
Det tidlige blomster- og bærfald må ikke forveksles med den "drysning" af bær, som, afhængig af sorten, sker, når modningen skrider frem (bilag 4-13).
Bilag 4-13. Virkning af planteafstand på vækst- og blomsterudvikling hos solbær. 2. vækstår (Larsen 1968).
Afstand, cm: |
75 x 75 |
50 x 50 |
25 x 25 |
15 x 15 |
Bladarealindeks |
4,6 |
6,8 |
7,8 |
8,0 |
m skud pr. m² |
6 |
12 |
18 |
28 |
Blomsterantal pr. klase |
21 |
22 |
22 |
15 |
% frugtsætning |
41 |
25 |
19 |
21 |
gram pr. bær |
1,36 |
1,27 |
1,00 |
0,86 |
ton pr. ha |
21,9 |
25,0 |
23,3 |
20,7 |
4.4. Frø- og frugtudvikling. Parthenocarpi. Vækststoffer
Udvikling af frø kan have betydning for frugtsætningen (såvel som for frugtstørrelse og frugtform, se afsnittene 6.11, 6.12). Hos fx æble og solbær vil en mere effektiv bestøvning give flere frø pr. frugt og en større frugtsætning. Ved afskæring af et stigende antal grifler inden bestøvning fås færre frø og øget frugtfald hos æble. Æbler, som falder af ved junifald, har færre frø end de frugter, som bliver siddende.
Frøenes rolle hænger muligvis sammen med en direkte eller induceret vækststofproduktion. Der produceres både auxiner, gibberelliner og cytokininer i frø (bilagene 4-14, 4-15), og det periodiske forløb er sat i relation til faser i frugtsætning og frugtfald (bilag 4-14). Hos arter med mange frø er der også en sammenhæng mellem frøantal, vækststofproduktion og frugtstørrelse (se kap. 6).
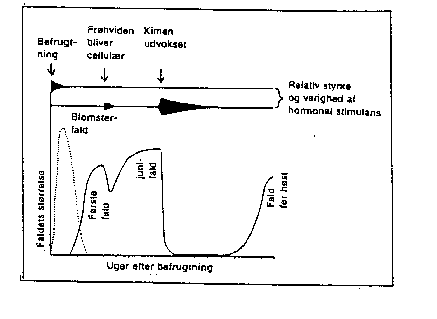
Bilag 4-14. Skematisk fremstilling af sammenhænge mellem frøudvikling, auxin-produktion og frugtfald hos æble. Efter Grauslund (1978); (Luckwill 1953).
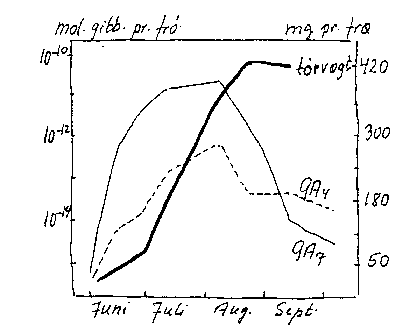
Bilag 4-15. Sæsonmæssig ændring i æblefrøs tørstof- og gibberellinindhold. Efter Sinska et al. (1973).
Det er dog ikke i alle tilfælde, at man kan se en sammenhæng mellem frø og frugtudvikling. Hos triploide æblesorter sker frugtsætning til trods for et relativt ringe frøantal pr. frugt. I afsnit 4.1 er omtalt, at en vis frugtsætning kan ske ved selvbestøvning af fremmedbefrugtere, hvor der så udvikles frugter med få frø eller helt parthenocarpe frugter. En sådan frugtudvikling sker dog kun under gunstige forhold (ringe konkurrence i træet), hvor der ikke samtidigt findes frugter med god frøudvikling i træet.
En vis tendens til parthenocarp frugtudvikling forekommer hos æble og især pære, der er en stor sortsvariation (Karnatz 1963). Denne tendens kan fremmes ved vækststoftilførsel. Dette viser igen vækststoffers rolle i frugtsætningsprocessen. Der er lavet mange forsøg med vækststoftilførsel, gennemgående med en betydelig variation i resultaterne (Schwabe & Mills 1981). Gibberelliner (GA3 = gibberellinsyre) er mest undersøgt. Hos nogle pæresorter kan der opnås virkning ved sprøjtning med 10-25 ppm ved 30-80% åben blomst. Koncentrationen skal øges i takt med den effekt, man ønsker, dog afhængig af sortens følsomhed (tendens til misdannede frugter).
Blandinger af auxiner, gibberelliner og cytokininer er også undersøgt. Auxin + gibberellin alene synes dog at have givet lige så godt resultat og har bl.a. virket i et dansk forsøg på sødkirsebær (bilag 4-16). Det må bemærkes, at man normalt ikke får virkning, hvis sætningsniveauet i forvejen er højt, men kun hvis det er lavt, fx på grund af delvis frostskade på blomsterne. Men det er jo også her, at en virkning er særlig ønskelig.
Bilag 4-16. Virkning af gibberellin- + auxin-tilførsel på frugtsætningen hos sødkirsebær (Rasmussen & Grauslund 1983).
% initial |
% endelig |
|
1981: Ubehandlet |
10 |
0,1 |
---GA3 + NOXA |
17 |
11 |
---GA3 + NAA |
17 |
13 |
1982: Ubehandlet |
69 |
51 |
---GA3 + NAA |
80 |
41 |
Forsøg har vist, at AVG (aminoethoxyvinylglycin), som hæmmer biosyntesen af ethylen, også kan øge frugtsætningen, formentlig ved at reducere det "naturlige" frugtfald. Ethylen fremmer abscission og findes naturligt i planten (se afsnit 9.2). Både ved tilførsel om efteråret samt før og under blomstringen kan AVG øge sætningen.
Tilførsel af vækststoffer med henblik på at øge frugtsætning er kun anvendt lidt i praksis, nok fordi behandlingen er relativt dyr, og virkningen kan være usikker (varierende) selv under gunstige forhold. Desuden kan der være uheldige bivirkninger; gibberellinbehandlede pærer kan således være langagtige, undertiden direkte misformede.
Følgende oversigt viser, at frugtsætning påvirkes af stort set de samme faktorer, som blomsterknopdannelse og blomsterudvikling.
Blomsterknopdannelse, hhv. blomsterudvikling, hæmmes af: |
Udvikling af æganlæg, EPP, (initial) sætning hæmmes af: |
Junifald øges af (især hos storfrugtede arter): |
Stor bæring samme år (især storfrugtede arter) |
(stor bæring året før, storfrugtede arter) |
Stor bæring samme år |
Stor skudvækst samme år |
Stor skudvækst året før |
Stor skudvækst samme år |
Lysmangel samme år |
Lysmangel året før |
(Lysmangel samme år) |
N-mangel |
N-mangel |
Vand (N) mangel |
Dårlig forsyning af knopper forår |
Dårlig forsyning af knopper forår |
Dårlig forsyning af frugter |
Speciel blomsterposition (især Ribes, Rubus, jordbær) |
Speciel blomsterposition (især Ribes, Rubus, jordbær) |
(Speciel blomsterposition, drysning solbær) |
Frugtsætning fremmes direkte af: God forenelighed med pollen; God bestøvning (rigelig pollen, god pollenoverførsel); Visse vækststoffer, især ved lavt sætningsniveau.
Junifald fremmes direkte af: Frugtudtyndingsmidler.
Vasilakatis, M. & I.C. Porlingis 1985. Effect of temperature on pollen germination, pollen tube growth, effective pollination period, and fruit set of pear. Hort. Science 20:733-735.
Williams, R.R. & M. Maier 1977. Pseudocompatibility after self-pollination of the apple Cox's Orange Pippin. J. Hort. Sci. 52:475-483.
Karnatz, A. 1963. Untersuchungen über das natürliche Ausmass der Parthenocarpie bei Kernobstsorten. Der Züchter 33:249-259.
Hansen, P. 1989. Source-sink relations in fruits. V. Pollination, fruit set, seed number, and fruit growth in apple cv. 'Summerred'. Gartenbauwiss. 54:129-132.
Kühn, B.F. 1986. Bestøvning af hyld. Statens Planteavslforsøg meddelelse w. 1870.
Williams, R.R. & D. Wilson 1970. Towards regulated cropping. London. 61 pp.
Hansen, P. 1980. Virkning af skudvækst, bæring og blomstermængde på frugtsætningen hos æbletræer. Tidsskr. Planteavl 84:499-505.
Stösser, R. & S.F. Anvari 1990. Über die Lebensdauer von Sammenlagen in Beziehung zum Fruchtansetz beim Steinobst. Erw. Obstbau 32:134-137.
Stösser, R. & S.F. Anvari 1982. On the senescence of ovules in cherries. Scient. Hort. 16:29-38.
Jackson, J.E. & P.J.C. Hamer 1980. The causes of year-to-year variation in the average yields of Cox's Orange Pippin apple in England. J. Hort. Sci. 55:149-156.
Dalbro, S. 1966. De små blade hos æbletræer. Horticultura 20:4-6.
Hansen, P. 1981. Bestøvning og frugtsætning hos 'Stevnsbær'. Tidsskr. Planteavl 85: 411-419.
Kühn, B.F. 1988. Examination of reasons for poor fruit set in the sour cherry cultivar 'Stevnsbær' by means of fluorescens microscopy. Tidsskr. Planteavl 92:169-174.
Lech, W. & K. Tylus 1983. Pollination, fertilization and fruit setting of some sour cherry varieties. Acta Hort. 139:33-39.
Bangerth, F. 1989. Dominance among fruits/sinks and the search for a correlative signal. Physiol.Plant. 76:608-614.
Habdas, H., L.S. Jankiewics & B. Borkowska 1982. Changes in anatomical structure of apple fruitlet pedicels preceding June drop. Acta Bot. 35:11-23.
Berüber, J. & P. Droz 1991. Studies on loacting the signal for fruit abseission in the apple tree. Scient.Hort. 46:201-214.
Grauslund, J. 1978. Effects of temperature, shoot-tipping, and carbaryl on fruit set of apples. Acta Hort. 80:207-211.
Polomski, R.F., J. Barden, R.E. Byers &D.D. Wolf 1988. Apple fruit nonstructural carbohydrates and abscission as influenced by shade and terbacil. J. Amer. Soc. Hort. Sci. 113:506-511.
Schanz, R. & R. Stösser 1989. Chromosomenzahl im Endosperm abortierender Steinobstfrüchte. Gartenbauwiss. 54:264-268.
Daugaard, H. 1981. Faktorer af betydning for blomster- og bærfald hos solbær. En litteraturoversigt. Beretning S 1559 fra Statens Planteavlsforsøg. 45 pp.
Kühn, B.F. 1988. Frugtfald i solbær. 1. Eksterne forhold. 2. Virkning af vækstregulerende stoffer. Tidsskr. Planteavl 92:59-67, 69-73.
Toldam-Andersen, T.B. & P. Hansen 1983. Growth and development in black currants (Ribes nigrum) I. Effects of light and leaf-shoot removals on growth distribution and fruit drop. Acta Hort. 352:237-246.
Hansen, P. & J. Ostermann 1988. Source-sink relations in fruits. II. Fruit growth and composition in black currants (Ribes nigrum L.). Gartenbauwiss. 53:155-159.
Larsen, F. 1968. Spacing trial with black currant. Roy. Vet. and Agric. Coll. Yearbook 1968: 49-58.
Wertheim, S.J. 1971. The drop of flowers and fruits in apple, with special reference to the June drop of Cox's Orange Pippin and its control with growth regulators. Meded. Landbouwhogeschool Wageningen 71-17.
Grauslund, J. 1978. Frugtudtynding IV. En oversigt over kemisk frugtudtynding. Tidsskr. Planteavl 82:521-539.
Sinska, I., M.J. Grochowska & S. Lewak 1973. Changes in the endogenous gibberellins contents in immature apple seeds. Bull. l'Academie Pol. Sci. V:26.
Schwabe, W.W. & J.J. Mills 1981. Hormones and parthenocarpic fruit set. A literature survey. Hort. Abstracts 51:661-698.
Goldwin, G.K. 1984. Factors affecting hormone-assisted setting of Cox's apple. Acta Hort. 149:161-171.
Rasmussen, K. & J. Grauslund 1983. Vækstregulatorer til frugttræer. VI. Virkning af gibberellin og auxin på frugtsætning og frugtkvalitet i kirsebær. Tidsskr. Planteavl 87:605-616.
Child, R.D. & R.R. Williams 1983. The effects of the interaction of aminoethoxyvinylglycerin (AVG) with mixtures of growth regulators on fruit set and shape in Cox's Orange Pippin apple. J. Hort. Sci. 58:365-369.